Eye like having lots of opsins – part 3
- May 5, 2020
- 9 min read
Updated: May 7, 2020

In Jan 1915, not long after the Great War began, Mildred Hoge, a newly minted Ph.D. geneticist from Columbia University now working as a zoology instructor at Indiana University, reported in The American Naturalist that she had found a new fruit fly gene which she named eyeless, after the phenotype (or form) of the mutation which lacked eyes.
The important thing is that not only did Hoge find this rare mutant, but she bred the fragile and short-lived eyeless fly, determined quantitatively how the mutation was inherited, and found that the gene controlling this mutation was located on the 4th of four fly chromosomes. Hoge found this dramatic physical effect whose cause she traced to a specific location (a chromosome) in the cell.

In December of 1919 Mildred married Aute Richards, who became head of the University of Oklahoma’s department of Zoology soon after they married. He eventually had a building (built during the depths of the Great Depression as a WPA project) named after him. Meanwhile, Mildred, supposedly due to nepotism rules, was unable to get a position at the university. Finally, just a year before she retired in 1948, she secured an associate professorship. A year!
In 1925, Mildred Hoge Richards published a detailed anatomical comparison of eyeless flies and normal flies, spurred by a question posed by Thomas Hunt Morgan, the Nobel Prize winning geneticist who set up the famous Fly Room at Columbia University. (I assume that Hoge got her Ph.D. working for Morgan at Columbia University, though I can’t find any specific references. My assumption is based on the fact that she got her Ph.D. in genetics from Columbia University in 1914 and was obviously well trained in fly genetics at a time when Morgan’s Fly Room was the only place to get such training. Morgan references several of Hoge’s papers including her report of the eyeless gene in his seminal book “The Mechanism of Mendelian Heredity”. And she clearly had a collaborative relationship with him as her 1925 paper attests.) Hoge Richards, responding to Morgan’s query, reported that mutation of the gene called eyeless appeared to have variable effects from reduced to eliminated eyes. In reduced eyes, the optic tract (the bundle of nerves that carry visuals signal from the light-sensing cells to the brain) were fully present. In flies that lacked eyes, the outermost optical bundle (called the outer ganglion, closest to the where the eyes should have been) was gone. The two inner bundles of optical nerves (middle and inner ganglion) closest to the brain remained in the totally eyeless flies. This seemed to establish the physical scope of influence of the eyeless gene. We’ll talk shortly about what this eyeless gene does.
In the early 1940s, Herman B. Chase at the University of Illinois, Urbana, published a series of genetics papers on the inheritance and variability of the eyeless phenotype in mice and proposed the symbol ey for the mouse gene responsible, after the fruit fly nomenclature for its own eyeless gene.
In the many decades since T. H. Morgan and his disciples like Mildred Hoge first used the fruit fly (Drosophila melanogaster) to probe the mechanism of Mendelian inheritance, the fly had become one of the workhorses of genetics. Mutant strains, such as the eyeless strain characterized and developed by Hoge, had become the foundation for genetics research. In earlier posts I talked about how the basic machinery in the simplest organisms (like the single celled yeast that I use in my research) are powerful models for understanding those same functions in more complex organisms like humans… the same is true for studying flies as you’ll see in a bit. But yeast do have limitations being single-celled. Flies, however, like humans, are metazoans (multicellular), are bilaterally symmetric (left and right sides appear mirrored), are built on a segmented body plan (as I discussed in a previous post), and, relevant to our current topic, have eyes.
The power of Drosophila genetics made it ideal for the groundbreaking (and back-breaking) research in the 1970-80s by Christiane Nusslein-Volhard and Eric Wieschaus which led to their discovery of many of the important genes needed for embryonic development in flies (and as we later found, humans):
Eric Wieschaus was finishing a Ph.D. in the lab of Walter Gehring, and briefly met Nusslein-Volhard when she came to Gehring’s lab to begin a post-doc. We’ll see more of Gehring shortly. Despite their passing-ships acquaintance, Wieschaus and Nusslein-Volhard discovered a complementary partnership and began collaborating on and off for over a decade, screening thousands of embryonic fly mutants for developmental defects and painstakingly characterizing them. Many of the genes they found in the fly were later found to be highly conserved Hox genes that operate in human development as well (something I posted about before – remember the creepy hand x-ray?). This effort ultimately led to the Nobel Prize for Nusslein-Volhard and Weischaus, awarded in 1995.
In the mid 1980s Markus Noll’s lab, then in Basel, Switzerland, published a paper in Cell showing that, within some of the fly development genes found by Nusslein-Volhard and Weischaus, important domains (or parts) were conserved and used repeatedly in several related genes. One of the domains in these developmental genes was given the name of paired box, which ultimately became abbreviated by geneticists as Pax. What this paired box domain did was not known, only that it was conserved and shared among closely related fly developmental genes.
In 1989, Noll’s lab published another paper showing that these Pax-type genes exhibiting a paired-box domain were present not only in fruit flies, but in a wide range of animals from nematodes to humans.

It seems like we’ve meandered far off the path of our story on eyes by talking about the discovery of developmental and Pax genes in diverse species, until around 1991 when a human gene associated with aniridia (a human genetic condition characterized by loss or reduction of the iris) was shown to encode a protein containing a paired box domain. It was a Pax gene.
This gene associated with eye development was eventually named Pax6, and the name was applied to mouse and other vertebrates with genes that were homologous (or related) to the human one. We later learned that Pax6 is so strongly conserved among diverse species that between mouse and human Pax6 proteins, there is only one difference out of 422 amino acids.
Then in 1994 Walter Gehring’s lab (who hired Nusslein-Volhard and Wieschaus and fatefully introduced them) published a paper in Science which showed that the gene ey (eyeless) in fruit flies was homologous to Pax6 in mice, rats, quail, zebrafish, and human.
A crucial question remained – these genes have historically been identified by (and named as) the mutant form of the organism when the gene is disabled (mutate a gene in drosophila, and the fly becomes eyeless, so we name the gene eyeless). But you can disable many things in a car and make it not operate – the question is whether the thing you disabled actually controls and regulates the car. Similarly, the question remained after all the decades of research briefly summarized above: do we know for sure that this eyeless (or Pax6) gene is actually used to develop eyes in these organisms? So far, we have only disabled the gene, and the car doesn’t operate, or the eye doesn’t develop correctly. We don’t know.

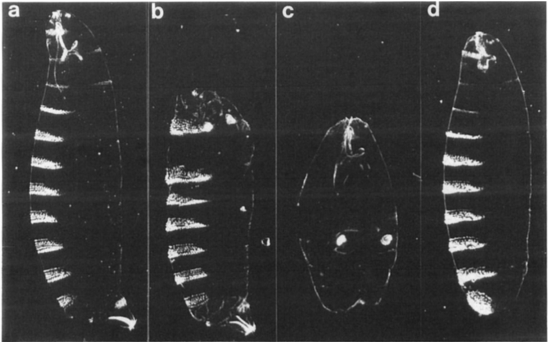
One method biologists use to tell if a gene does what we hypothesize is to force the normal gene to function in a location where it is not normally expressed. Biology is loaded with jargon like any specialized field, and the fancy word used for this kind of experiment is “ectopic”… we want to express the Pax6/eyeless gene ectopically (some place other than the eye region) and see if the gene makes an eye outside its normal field of influence. If not then we conclude the gene does not specify eye development (or that something else is wrong in the experiment). If an eye develops then we conclude that indeed Pax6 is an eye development regulator (it's not so simple as this, but that's the basic idea). This Frankensteinian experiment is exactly what Gehring’s lab did and published in 1995 in Science, showing that by expressing Pax6 in ectopic locations such as the wings and legs, all the anatomical features (even the red color) of a fly eye developed.
This experiment established that the Pax6 gene did indeed control development of the eye. In the following years other labs repeated this result in more complex organisms such as vertebrates. We've now answered the question of whether Pax6 regulated eye development. But this was just a start, as the efforts to dissect the details of eye development accelerated… the next question was how Pax6 regulated eye development.
Czerny and Busslinger published a paper also in 1995 showing that Pax6 bound to a specific DNA sequence, and that Pax6 formed a dimer (a pair) and bound to an inverted repeat of the sequence TAAT with the repeats separated by three nucleotides. So a Pax6 DNA binding site might look like TAATxxxATTA (where x = A, G, C or T).
Interestingly, Czerny and Busslinger used sea urchin Pax6 which they showed was very similar to Pax6 from mouse and fruit fly. They also pointed out amino acids that were identical across all these species, and further narrowed it down to three amino acids that controlled the specific binding of Pax6 to its TAATxxxATTA DNA binding site.

In 1999 Xu et al published the detailed structure of the human Pax6 paired domain bound to DNA, obtained using X-ray crystallography. This structure showed at an atomic level of resolution how the paired domain is able to specifically bind to a particular DNA sequence, and therefore regulate formation of a structure at a defined location like the eye.
In 2001, Marquardt et al demonstrated that Pax6 is the master switch that controls the development of all the retinal cells in vertebrates.
We should pause here to recap and see where we are even though we are almost done. We learned from very early work on eyeless mutations in animals as diverse as fruit flies and mice that there was a gene that appeared to control development of the eye. This was identified as the now commonly called Pax6 gene which is known to be directly responsible for the development of eye-specific cells such as the diverse cells in the retina. Pax6 is a highly conserved gene and appears to retain this common function of specifying the eye despite the amazing diversity of species and eye plans. Furthermore, we learned that the protein encoded by the Pax6 gene binds directly to a specific sequence in DNA, from which it acts as a master switch to control the development of retinal cells. (There is much more of course, we know things like humans have a family of Pax genes with at least nine members, and that Pax6 has more jobs than specifying the eye, much like our Hox genes do more than just regulate the development of our segmented - or vertebral - body plan.)
So then, what is this thing, this Pax6? Molecular biologists call proteins like Pax6 a transcription factor, but that bit of jargon basically means it acts like a molecular network switch. Let’s tease apart what that actually means. We know genes store the information needed to make a protein. But to make a protein, the first step is to transcribe the data encoded in DNA into a message in the form of RNA (which will exit the nucleus into the cytoplasm where it will be translated into protein). But for the DNA to be transcribed into RNA, we need a large molecular machine called an RNA polymerase to read the DNA and create a single copy of RNA each time it scans along the gene. For that RNA polymerase to know when and where to start, transcription factors bind to specified regions around the gene to be transcribed… the transcription factors signal to the RNA polymerase where to bind, and when to start transcribing.
Most often, most of the genes in each cell in our body are switched off (and there are many layers of switches and other mechanisms that ensure that genes intended to be off remain mostly off).
A master regulator, like Pax6, is a transcription factor that turns on (or off) the genes for other transcription factors which, in turn, turn on (or off) genes for yet other transcription factors… in a cascading sequence of switches. Sometimes these switches turn themselves on or off – or they can operate on switches upstream of themselves like the master switch Pax6 – creating a feedback loop. Eventually, some of those switches or transcription factors (usually a combination of them) turn on genes needed to make a retina cell, the genes to make the proteins that make that specific cell function correctly (like rhodopsin proteins required for a rod cell to detect a photon). All this layered, networked, feedbacked switching of transcription factors seems complicated – but recall that we are trying to carefully control which cells turn into eye cells, and nearby, which cells turn into nose cells. You don’t want to get those confused (though it does happen sometimes).
So now we know additionally that Pax6 is a transcriptional regulator controlling when genes necessary for eye development are active. The genes that Pax6 switches on or off determine whether the cell becomes an eye cell, such as those in the retina. Some of the important cells in the retina directly involved in the detection of light include rod and cone photoreceptor cells as we posted recently. Furthermore, one of the proteins that determine the function of a photoreceptor cell are rhodopsins.
In 1997 and 2001, the lab of Claude Desplan published papers showing that Pax6/eyeless in Drosophila regulates expression of rhodopsin 1, one of the light-sensing molecules essential for vision in a wide range of organisms from flies to humans that we discussed in a previous post.
There! We’ve finally meandered along and linked the Pax6 genes responsible for eyeless fly mutants discovered at the beginning of the last century, to the rhodopsin molecules which we discussed earlier as directly responsible for vision in almost anything with eyes. I think Mildred Hoge would have been proud to be an essential part of this story. Geewiz indeed!



Comments